在国家自然科学基金等项目资助下,华南农业大学农学院教授陈乐天/刘耀光团队研究揭示了水稻Sa座位三基因“杀手-保镖”系统的分子遗传基础,首次阐明植物杂种不育响应环境温度的全新分子机理。12月11日,相关成果在线发表于《分子植物》(Molecular Plant)。
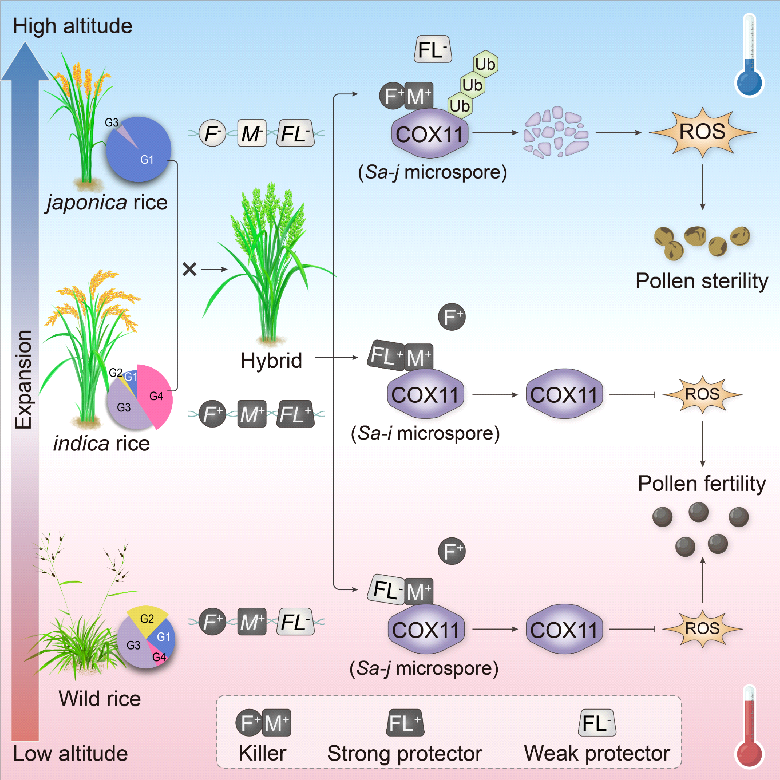 Sa在水稻亚种间分化及其介导的杂种不育的机制模型。研究团队供图
Sa在水稻亚种间分化及其介导的杂种不育的机制模型。研究团队供图
生殖隔离是物种形成的关键环节,它通过阻止不同种群间的基因交流,推动物种分化。杂种不育作为典型的合子后生殖隔离系统,成为制约农作物杂种优势利用的一大障碍。过去10多年间,多个植物(尤其是水稻)的杂种不育位点被成功克隆,但植物杂种不育受温度等环境因素调控的分子机制始终未被破解。
研究团队前期在水稻籼粳杂种不育Sa复合座位中,鉴定出两个紧密相邻的关键基因SaF和SaM。其中,SaF编码F-box蛋白,籼稻的SaF+与粳稻的SaF-仅有一个氨基酸的差异;籼稻的SaM+编码具有完整SPL-RING结构域的蛋白,而粳稻的SaM-编码的则是结构域缺失的截短蛋白。后续研究中,团队发现Sa座位介导的F1杂种花粉育性受温度影响显著:在较低温条件(23-29℃,如晚季)下,花粉育性约为50%;而在高温条件(30-35℃,如早/中季)下,花粉育性提升至75%左右,这表明高温能够缓解远缘杂交水稻的杂种不育。这一现象背后隐藏着两个关键科学问题:一是,Sa座位引发花粉败育的分子机制是什么?二是,杂种花粉育性为何在高温下不降反升?为此,团队展开了长达10多年的深入研究,最终揭开了其中的奥秘。
通过遗传群体分析,团队在Sa座位鉴定出一个与SaF、SaM相邻的新基因SaFL,该基因决定杂种不育的温度敏感性。SaFL编码F-box类似蛋白,籼粳稻的SaFL蛋白仅存在2个氨基酸的差异。互补实验结果显示,籼稻SaFL+可恢复杂种F1花粉育性;在F1中敲除SaFL+,mF1突变体在低温下全不育,高温下恢复为半不育;在F1中敲除SaFL-,温敏性消失,mF1花粉在高低温下均表现为半不育。这些结果充分证明,SaFL+是强功能的花粉保护(保镖)基因,SaFL-则仅在高温条件下发挥弱功能的花粉保护作用。遗传分析进一步表明,来自籼稻的SaF+和SaM+构成了Sa座位不可或缺的杀手系统。蛋白互作及竞争实验证实,SaM+与SaF+、SaFL+/-均存在互作关系,其中SaM+与SaF+互作的杀手系统不仅受高温抑制,还会因SaFL+/-竞争性结合SaM+而遭到破坏。研究还揭示,Sa杀手复合体具有线粒体E3连接酶功能,可靶向线粒体活性氧清除蛋白COX11,介导其泛素化和26S蛋白酶体降解,进而引起活性氧过量积累,最终导致花粉不育。
进一步的单倍型分化分析显示,Sa三基因复合座位起源于非AA基因组的远缘野生稻,随后向AA基因组野生稻扩散。在栽培水稻驯化及从热带向温带扩张的过程中,携带杀手基因和弱保护者的单倍型G2在野生稻中持续存在,但在温带地区的栽培稻中逐渐被淘汰,籼型和粳型逐渐成为(亚)热带和温带地区的主流单倍型。由此推测,Sa座位的温敏特性可能是驱动水稻种群纬度适应和籼粳亚种形成的关键因素。最后,团队提出了Sa座位介导杂种不育及其演化的分子模型,为利用生物技术(转基因和基因编辑)克服远缘杂种不育、建立新型两系杂交制种技术提供了坚实的理论依据。
在水稻籼粳杂种雄性不育复合座位Sa中,SaF、SaM和SaFL共同构成一个温度敏感的花粉“杀手-保镖”系统。其中,籼稻的SaF+和SaM+形成复合体作为“杀手”,靶向线粒体活性氧清除蛋白COX11,促进其泛素化降解,导致活性氧过量积累,从而引发花粉不育。籼稻中的SaFL+是强功能的“保镖”,粳稻中的SaFL-仅在高温下发挥作用,为弱功能“保镖”。在籼粳杂种中,一半花粉携带籼稻强功能SaFL+,它能竞争性优先结合SaM+,破坏杀手复合体,保护花粉正常发育;另一半花粉携带弱功能SaFL-,其在高温下才发挥功能,使花粉呈现低温不育、高温可育的特性。
该研究从遗传、分子和进化三个层面展开,深入挖掘了三基因“杀手-保镖”系统响应环境高温调控水稻杂种不育的新机制,不仅深化了我们对生殖隔离机制的理解,也为杂种优势利用和杂交育种技术创新指明了方向。(来源:中国科学报 朱汉斌)
相关论文信息:https://doi.org/10.1016/j.molp.2025.12.008